
Poster Session D
Immunobiology
Robert Lochhead, PhD
Medical College of Wisconsin
GERMANTOWN, WI, United States
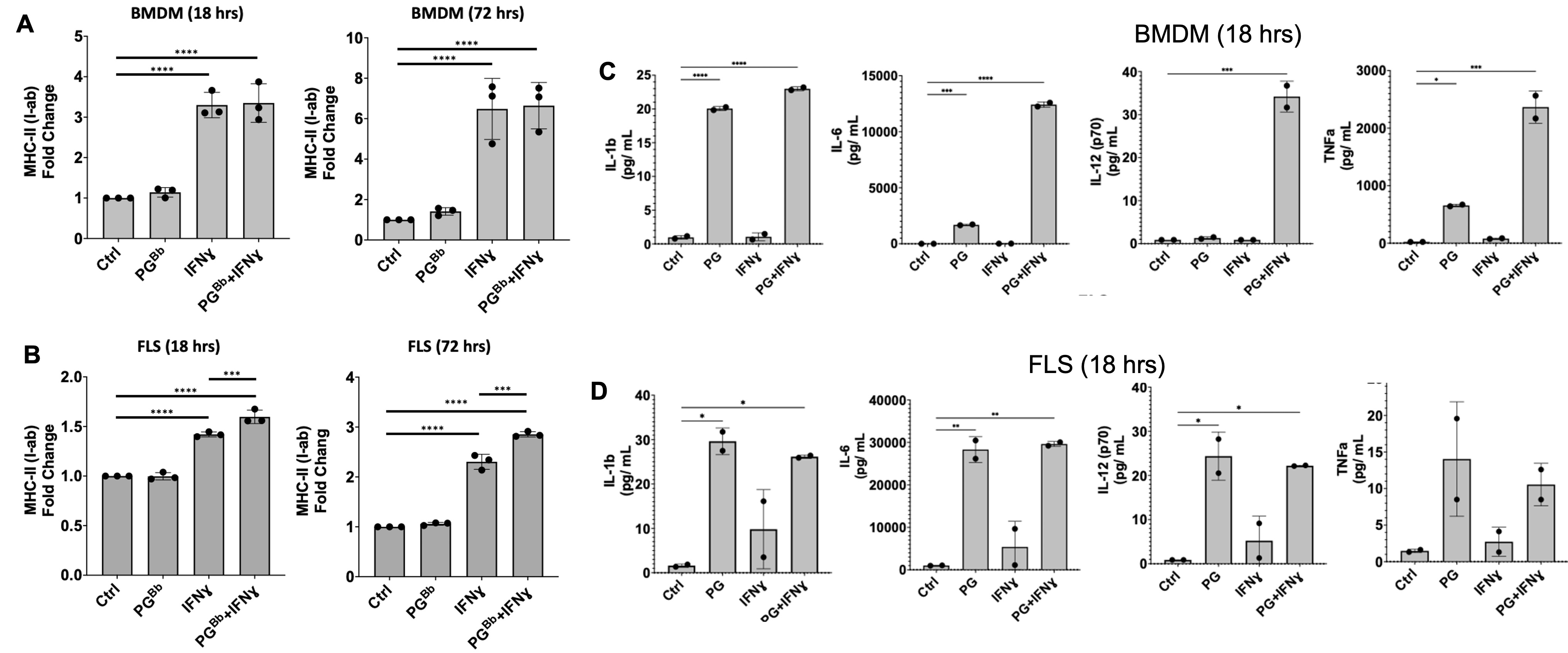 Figure 1: FLS immune effector phenotype in response to PG. B6 mouse BMDM and FLS surface expression of MHC-II (I-ab in mice) (left) and proinflammatory cytokine production (right) when stimulated with IFNɣ, PG, or both. (A) BMDM upregulated MHC-II surface expression in an IFNɣ-dependent manner, as expected. (B) FLS primed with IFNɣ upregulate MHC-II when stimulated with IFNɣ and B. burgdorferi PG for 18 hrs and this response was strengthened after prolonged exposure at 72 hrs. (C) BMDM and (D) FLS upregulation of proinflammatory cytokines in response to stimulation with PG muropeptides for 18 hrs. ***P < 0.0001, **P < 0.001, *P < 0.01
Figure 1: FLS immune effector phenotype in response to PG. B6 mouse BMDM and FLS surface expression of MHC-II (I-ab in mice) (left) and proinflammatory cytokine production (right) when stimulated with IFNɣ, PG, or both. (A) BMDM upregulated MHC-II surface expression in an IFNɣ-dependent manner, as expected. (B) FLS primed with IFNɣ upregulate MHC-II when stimulated with IFNɣ and B. burgdorferi PG for 18 hrs and this response was strengthened after prolonged exposure at 72 hrs. (C) BMDM and (D) FLS upregulation of proinflammatory cytokines in response to stimulation with PG muropeptides for 18 hrs. ***P < 0.0001, **P < 0.001, *P < 0.01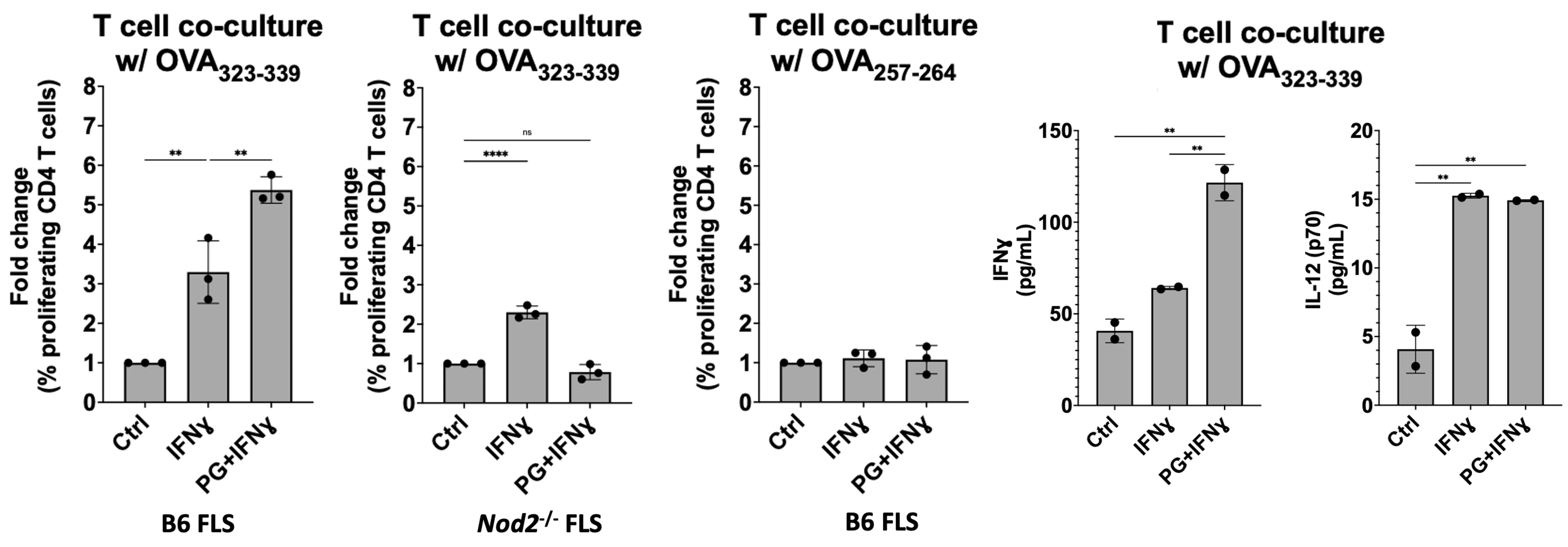 Figure 2: MHC-II+ FLS activate peptide-specific CD4+ T cell proliferation. IFNɣ-primed MHC-II+ B6 FLS were co-cultured with OT-II mouse T cells (genetically modified to have identical antigen-specific T cell receptors) and loaded with the cognate OVA323-339 peptide. T cell responses measured by flow cytometry and multiplex immunoassay. T cell co-culture with MHC-II+ FLS primed with IFNɣ induced naive OT-II mouse CD4+ T cell proliferation at a 3-fold increase compared to unstimulated FLS, which was enhanced to above 5-fold when co-cultured with FLS primed with both IFNɣ and PG. Negative controls using irrelevant OVA peptide or NOD2-deficient FLS failed to induce T cell proliferation.
Figure 2: MHC-II+ FLS activate peptide-specific CD4+ T cell proliferation. IFNɣ-primed MHC-II+ B6 FLS were co-cultured with OT-II mouse T cells (genetically modified to have identical antigen-specific T cell receptors) and loaded with the cognate OVA323-339 peptide. T cell responses measured by flow cytometry and multiplex immunoassay. T cell co-culture with MHC-II+ FLS primed with IFNɣ induced naive OT-II mouse CD4+ T cell proliferation at a 3-fold increase compared to unstimulated FLS, which was enhanced to above 5-fold when co-cultured with FLS primed with both IFNɣ and PG. Negative controls using irrelevant OVA peptide or NOD2-deficient FLS failed to induce T cell proliferation.