
Poster Session D
Immunobiology
Jie An, PhD
University of Washington
Seattle, WA, United States
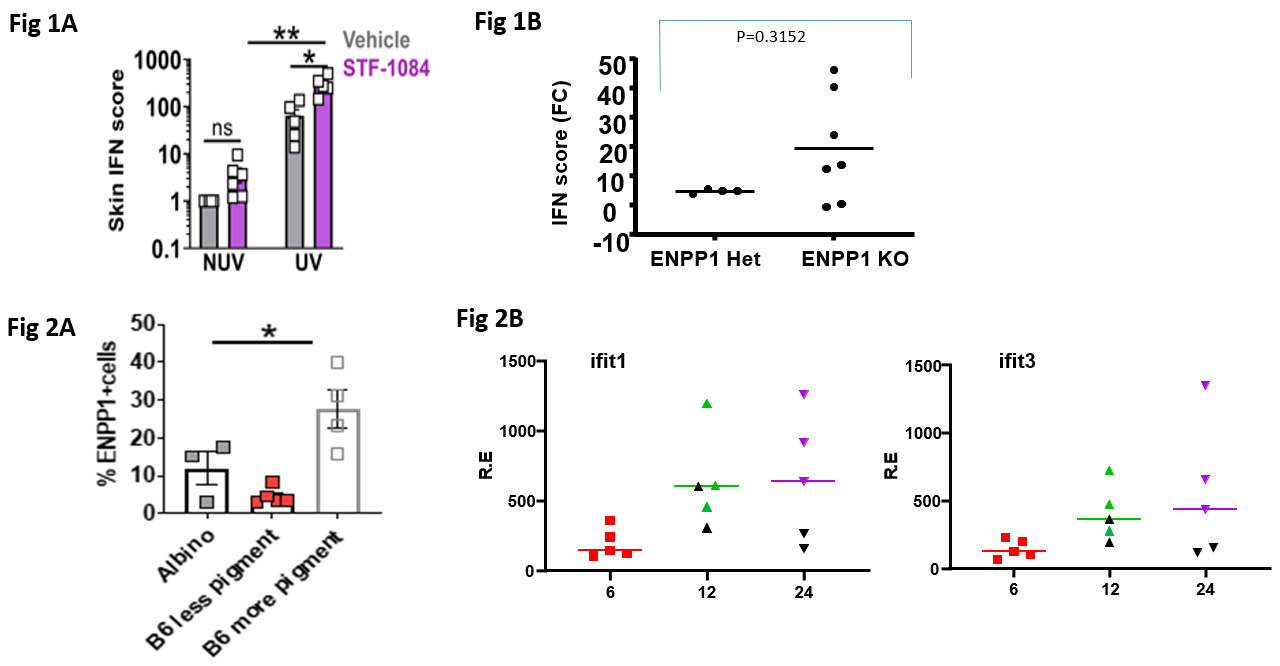 Figure 1: (A) Effect of local pre-application of the ENPP1 inhibitor, STF 1084, to mouse skin prior to UVB. (B) Effect of ENPP1 deficiency on UVB induced IFN responses. Figure 2. (A) Expression of ENPP1 on skin cells from pigmented and non-pigmented regions of skin. (B) Expression of ISGs (Ifit1 and Ifit3) in non-pigmented (green or purple symbols) compared to pigmented regions (black symbols) of skin following UVB exposure.
Figure 1: (A) Effect of local pre-application of the ENPP1 inhibitor, STF 1084, to mouse skin prior to UVB. (B) Effect of ENPP1 deficiency on UVB induced IFN responses. Figure 2. (A) Expression of ENPP1 on skin cells from pigmented and non-pigmented regions of skin. (B) Expression of ISGs (Ifit1 and Ifit3) in non-pigmented (green or purple symbols) compared to pigmented regions (black symbols) of skin following UVB exposure.